Como a diminuição da concentração extracelular de ions K+ (sem ocorrer alteração intracelular do mesmo) devido a urina, afeta o potencial de repouso?
fisiologia humana
💡 3 Respostas
Stephany Ellen
Pontos Principais:
-
Um neurônio em repouso (sem sinalização) tem uma voltagem em sua membrana chamada de potencial de repouso da membrana, ou simplesmente potencial de repouso.
-
O potencial de repouso é determinado pelos gradientes de concentração de íons na membrana e através da sua permeabilidade para cada íon.
-
Em um neurônio em repouso, existem gradientes de concentração na membrana para \text {Na}^+Na+N, a, start superscript, plus, end superscript e \text K^+K+K, start superscript, plus, end superscript. Os íons se deslocam de acordo com seus gradiente através de canais, ocasionando uma separação de cargas que resulta no potencial de repouso.
-
A membrana é muito mais permeável ao \text K^+K+K, start superscript, plus, end superscript do que ao \text {Na}^+Na+N, a, start superscript, plus, end superscript, por isso o potencial de repouso é próximo ao potencial de equilíbrio do \text K^+K+K, start superscript, plus, end superscript (o potencial que seria gerado pelo \text K^+K+K, start superscript, plus, end superscript se ele fosse o único íon no sistema).
Introdução
Vamos supor que você tem um sapo morto. (Sim, é um pouco nojento, mas vamos imaginar por um momento). O que aconteceria se você aplicasse um estímulo elétrico no nervo que inerva a perna do sapo? Assustadoramente, ela se moveria!
O cientista italiano Luigi Galvani descobriu esse fato cômico por volta de 1700, por acidente, enquanto dissecava um sapo. Hoje, nós sabemos que a perna do sapo se move porque os neurônios (células nervosas) carregam informações por meio de sinais elétricos.
Como neurônios em um organismo vivo produzem sinais elétricos? Basicamente, os neurônios geram sinais elétricos por meio de breves e controladas mudanças na permeabilidade de suas membranas celulares a íons específicos (como \text{Na}^+Na+N, a, start superscript, plus, end superscript e \text K^+K+K, start superscript, plus, end superscript). Antes de aprendermos em detalhes como esses sinais são gerados, primeiro precisamos entender como funciona a permeabilidade de uma membrana em um neurônio em repouso (que não está enviando nem recebendo sinais elétricos). Neste artigo, veremos como um neurônio estabelece e mantém uma voltagem estável em sua membrana – isto é, um potencial de repouso da membrana.
O potencial de repouso da membrana
Imagine dois eletrodos, um colocado fora e outro dentro da membrana plasmática de uma célula viva. Se fizer isso, você poderá medir a diferença de potencial elétrico, ou voltagem, entre os eletrodos. Essa diferença de potencial elétrico é chamado potencial da membrana.
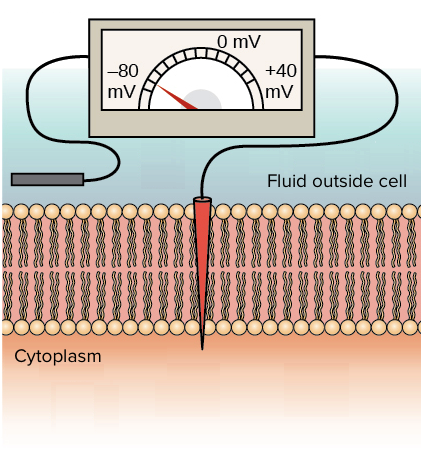
Diagrama de um voltímetro medindo o potencial da membrana. Um eletrodo se encontra fora da célula. O outro eletrodo está no interior da célula. O voltímetro mostra uma voltagem de -70 mV atravessando a membrana.
Assim como a distância, a diferença de potencial é medida em relação a um ponto de referência. No caso da distância, o ponto de referência pode ser uma cidade. Por exemplo, podemos dizer que Boston está a 300300300 \text{quilômetros}quiloˆmetros a nordeste, mas só se soubermos que nosso ponto de referência é a cidade de Nova York.
Para o potencial da membrana de uma célula, o ponto de referência é fora da célula. Na maioria dos neurônios em repouso, a diferença de potencial ao longo da membrana é em torno de 303030 a 909090 \text{mV}mVm, V (um \text{mV}mVm, V é 1/10001/10001, slash, 1000 de um volt), com o interior da célula mais negativo do que o exterior. Ou seja, os neurônios possuem um potencial de repouso da membrana (ou simplesmente, potencial de repouso) que varia de -30−30minus, 30 \text{mV}mVm, V até - 90−90minus, 90 \text{mV}mVm, V.
Por existir uma diferença de potencial na membrana celular, a membrana é dita polarizada.
-
Se o potencial da membrana se torna mais positivo do que está no potencial de repouso, então a membrana é dita despolarizada.
-
Se o potencial da membrana se torna mais negativo do que está no potencial de repouso, a membrana é dita hiperpolarizada.
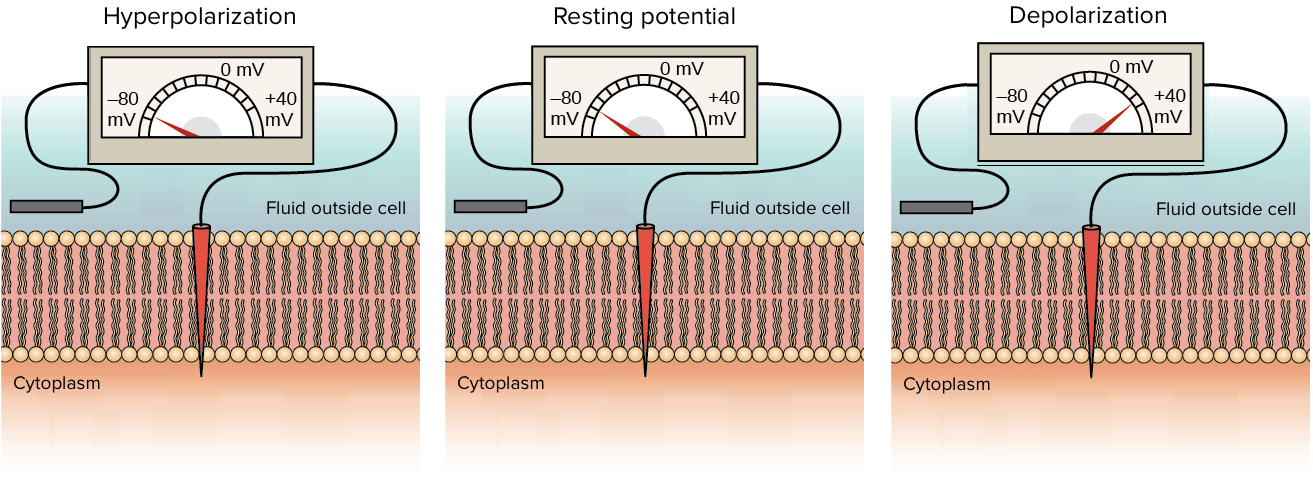
Diagramas de voltímetros com um eletrodo dentro da célula e um no fluido fora da célula. O primeiro voltímetro mostra hiperpolarização: lê-se -80 mV. O segundo voltímetro mostra o potencial de repouso: lê-se -70 mV. O terceiro voltímetro mostra despolarização: lê-se +40 mV.
Todos os sinais elétricos que os neurônios usam para comunicar são despolarizações ou hiperpolarizações do potencial de repouso da membrana.
De onde vem o potencial de repouso da membrana?
O potencial de repouso da membrana é determinado pela distribuição desigual de íons (partículas carregadas) entre o interior e o exterior da célula e pela permeabilidade da membrana diferenciada para diferentes tipos de íons.
Tipos de íons encontrados em neurônios
Os íons mais abundantes nos neurônios e no seu fluido circundante são:
- Íons carregados positivamente (cátions): Sódio (\text{Na}^+Na+N, a, start superscript, plus, end superscript) e potássio (\text{K}^+K+K, start superscript, plus, end superscript)
- Íons carregados negativamente (ânions): Cloreto (\text{Cl}^-Cl−C, l, start superscript, minus, end superscript) e ânions orgânicos
Na maioria dos neurônios, o \text{K}^+K+K, start superscript, plus, end superscript e os ânions orgânicos (tais como aqueles encontrados em proteínas e aminoácidos) estão presentes em maior concentração no interior da célula do que fora dela. Em contraste, o \text{Na}^+Na+N, a, start superscript, plus, end superscript e o \text{Cl}^-Cl−C, l, start superscript, minus, end superscript geralmente estão presentes em altas concentrações fora da célula. Isso significa que existe um gradiente de concentração estável através da membrana para todos os tipos mais abundantes de íon.
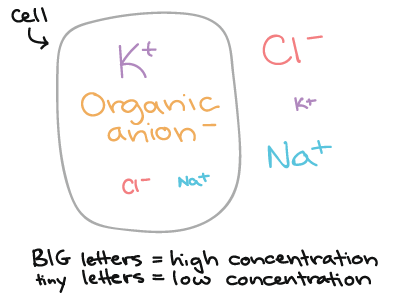
Este diagrama apresenta a concentração relativa de vários tipos de íons dentro e fora de um neurônio.
- K+ é mais concentrado no interior do que no exterior da célula.
- Ânions orgânicos são mais concentrados no interior da célula do que no exterior.
- Cl- é mais concentrado no exterior do que no interior da célula.
- Na+ é mais concentrado no exterior da célula do que no interior.
Como os íons atravessam a membrana
Por serem carregados, os íons não conseguem passam diretamente através de regiões lipídicas hidrofóbicas ("com medo de água") da membrana. Ao invés disso, eles usam canais proteicos especializados que fornecem um túnel hidrofílico ("amam água") que atravessa a membrana. Alguns canais, conhecidos como canais de vazamento, são abertos em neurônios em repouso. Outros estão fechados em neurônios em repouso, e só abrem em resposta a um sinal.
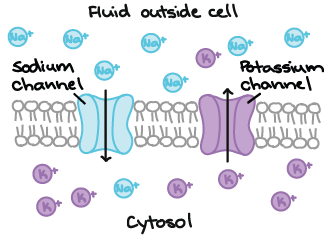
Canais iônicos. Os canais se estendem de um lado da membrana plasmática para o outro lado e apresentam um túnel no meio. O túnel permite que os íons atravessem a membrana. Um dos canais apresentados permite íons Na+ cruzarem e é um canal de sódio. O outro canal permite os íons K+ atravessarem e é o canal de potássio. Os canais simplesmente fornecem um caminho para que os íons atravessem a membrana, permitindo-os se deslocarem sob qualquer gradiente eletroquímico que possa existir. Os canais não movem ativamente os íons de um lado para o outro da membrana.
Alguns canais iônicos são altamente seletivos para um tipo de íon, mas outros permitem a passagem de vários tipos de íons. Canais iônicos que permitem principalmente a passagem de \text{K}^+K+K, start superscript, plus, end superscript são chamados canais de potássio e canais iônicos que permitem a passagem principalmente de \text{Na}^+Na+N, a, start superscript, plus, end superscript são chamados canais de sódio.
\text {Cl}^-C, l, start superscript, minus, end superscript
\text {Na}^+N, a, start superscript, plus, end superscript\text {K}^+K, start superscript, plus, end superscript
Nos neurônios, o potencial de repouso da membrana depende principalmente do movimento de \text {K}^+K+K, start superscript, plus, end superscript através dos canais vazantes de potássio. Vamos ver como isso funciona.
O que acontece se somente o \text K^+K+K, start superscript, plus, end superscript conseguir atravessar a membrana?
O potencial da membrana de um neurônio em repouso é determinado principalmente pelo movimento de íons \text K^+K+K, start superscript, plus, end superscript através da membrana. Então, vamos ter uma ideia de como o potencial da membrana funciona ao ver o que aconteceria em um caso onde somente \text K^+K+K, start superscript, plus, end superscript pode atravessar a membrana.
Começaremos com \text{K}^+K+K, start superscript, plus, end superscript em uma maior concentração dentro da célula do que no fluido circundante, assim como em um neurônio regular. (Outros íons também estão presentes, incluindo ânions que contrabalanceiam a carga positiva do \text K^+K+K, start superscript, plus, end superscript, mas eles não serão capazes de atravessar a membrana em nosso exemplo.)
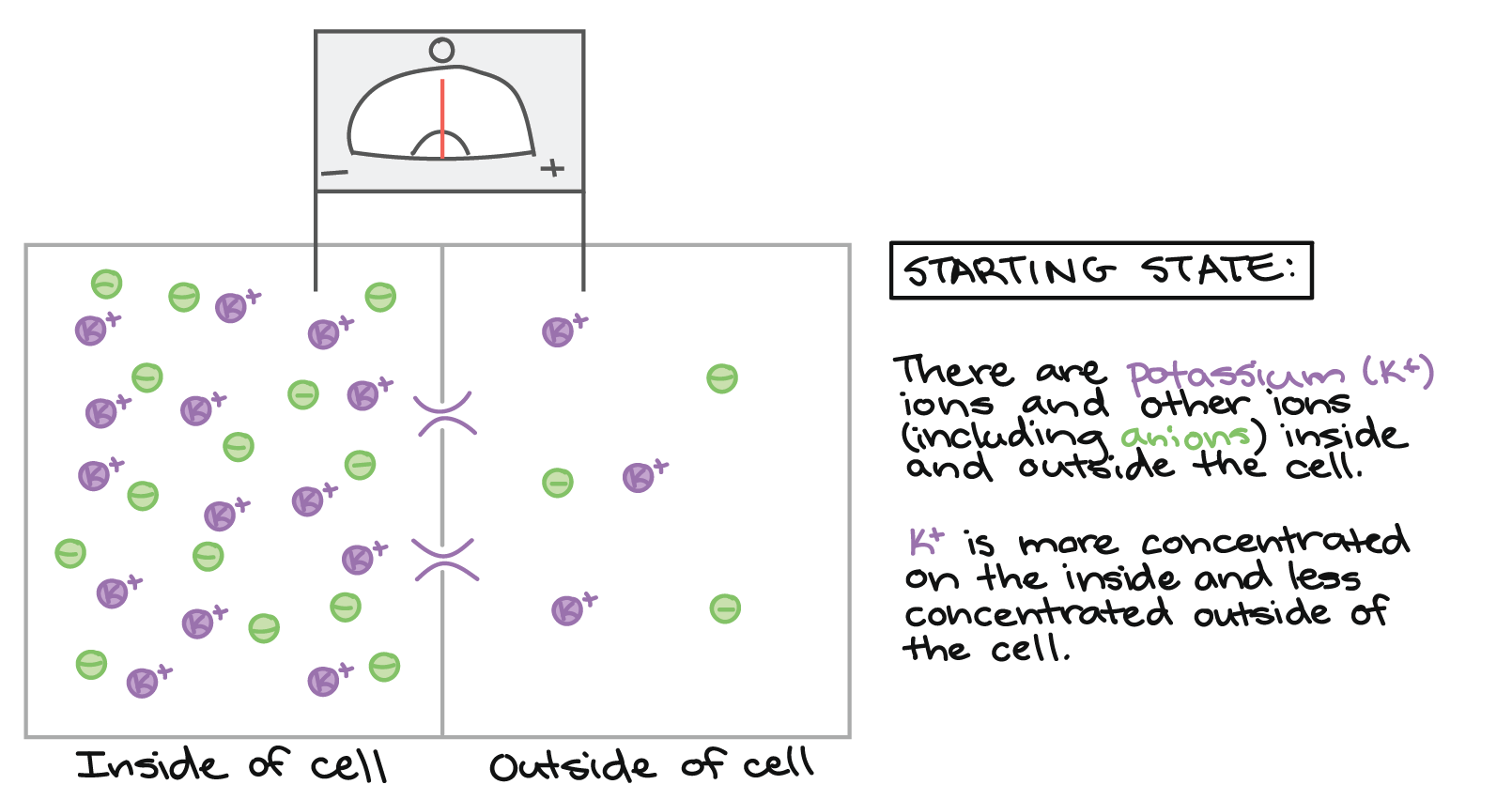
Estado inicial:
Voltagem zero através da membrana, como mensurado por um voltímetro com um eletrodo dentro e outro fora da célula. A parte de dentro da célula e a parte de fora da célula são separadas por uma membrana com canais de potássio, que estão inicialmente fechados. Há uma maior concentração de íons potássio dentro da célula do que fora dela. Cada íon potássio (de cada lado da membrana) é equilibrado por um ânion, então o sistema é neutro como um todo.
[Onde estão os íons Na+ e Cl-?]
\text K^+K, start superscript, plus, end superscript\text K^+K, start superscript, plus, end superscript\text {Cl}^-C, l, start superscript, minus, end superscript
\text {Cl}^-C, l, start superscript, minus, end superscript\text {Na}^+N, a, start superscript, plus, end superscript\text K^+K, start superscript, plus, end superscript
Se canais de potássio na membrana abrem, \text K^+K+K, start superscript, plus, end superscript começará a se mover de acordo com seu gradiente de concentração e para fora da célula. Toda vez que um íon \text{K}^+K+K, start superscript, plus, end superscript deixa a célula, o interior celular perde uma carga positiva. Por causa disso, um pequeno excesso de carga positiva se forma fora da membrana celular, e um pequeno excesso de carga negativa se forma dentro da célula. Isto é, o interior da célula se torna negativo em relação ao exterior, gerando uma diferença de potencial elétrico na membrana.
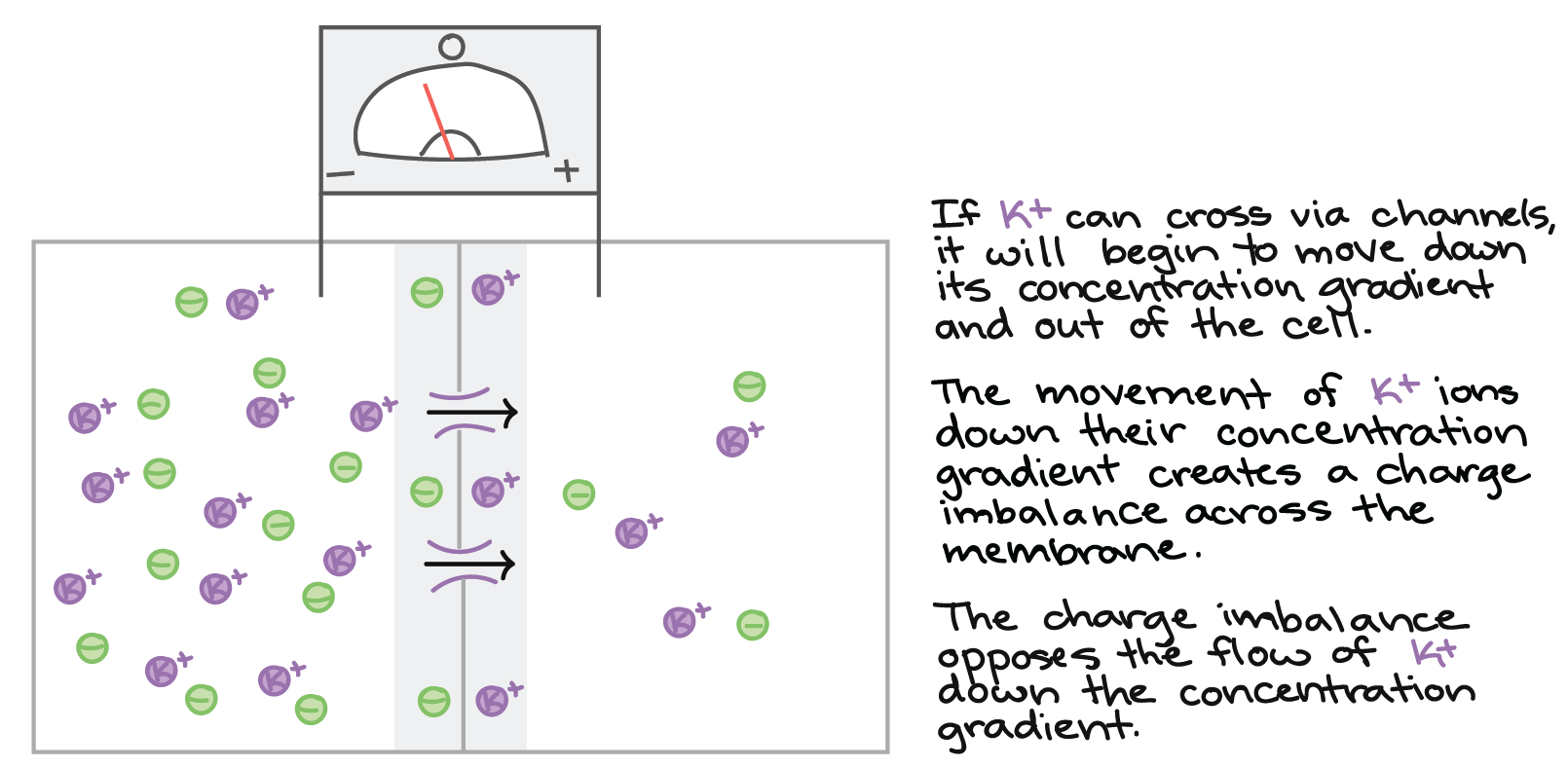
Sistema se movendo em direção ao equilíbrio:
Se o íon K+ pode atravessar por canais, ele começará a se mover pelo seu gradiente de concentração e para fora da célula. (Os canais são mostrados abrindo, o potássio é mostrado movendo-se do interior para o exterior da célula através dos canais).
O movimento de íons K+ pelo gradiente de concentração cria um desequilíbrio de carga através da membrana. (Os íons potássio que atravessaram do interior da célula para o exterior não se pareiam com ânions no exterior da célula. Eles se alinham ao longo da membrana no exterior, e os ânions não pareados que são deixados para trás no interior alinham-se ao longo da membrana na face interior. O voltímetro agora registra uma ligeira voltagem negativa).
O desequilíbrio da carga se opõe ao fluxo de K+ ao longo do gradiente de concentração.
Para íons (assim como para ímãs), cargas iguais se repelem e cargas diferentes se atraem. Logo, o estabelecimento da diferença de potencial elétrico através da membrana dificulta a saída dos íons \text K^+K+K, start superscript, plus, end superscript remanescentes da célula. Os íons \text K^+K+K, start superscript, plus, end superscript positivamente carregados serão atraídos pelas cargas negativas livres no interior da membrana da célula e repelidos pelas cargas positivas no exterior, oferecendo resistência ao seu movimento em direção ao gradiente de concentração. As forças elétricas e difusas que influenciam os movimentos de \text K^+K+K, start superscript, plus, end superscript através da membrana em conjunto formam o seu gradiente eletroquímico (o gradiente de energia potencial que determina em qual direção \text K^+K+K, start superscript, plus, end superscript fluirá espontaneamente).
No final, a diferença de potencial elétrico através da membrana da célula acumula-se a um nível alto o suficiente para que a força elétrica que conduz o \text{K}^+K+K, start superscript, plus, end superscript novamente para o interior da célula seja igual à força química conduzindo \text{K}^+K+K, start superscript, plus, end superscript para fora da célula. Quando a diferença de potencial através da membrana da célula alcança este ponto, não há nenhum movimento fluido de \text{K}^+K+K, start superscript, plus, end superscript em nenhuma direção e o sistema é considerado em estado de equilíbrio. Toda vez que um \text{K}^+K+K, start superscript, plus, end superscript deixa a célula, outro \text{K}^+K+K, start superscript, plus, end superscript entra nela.
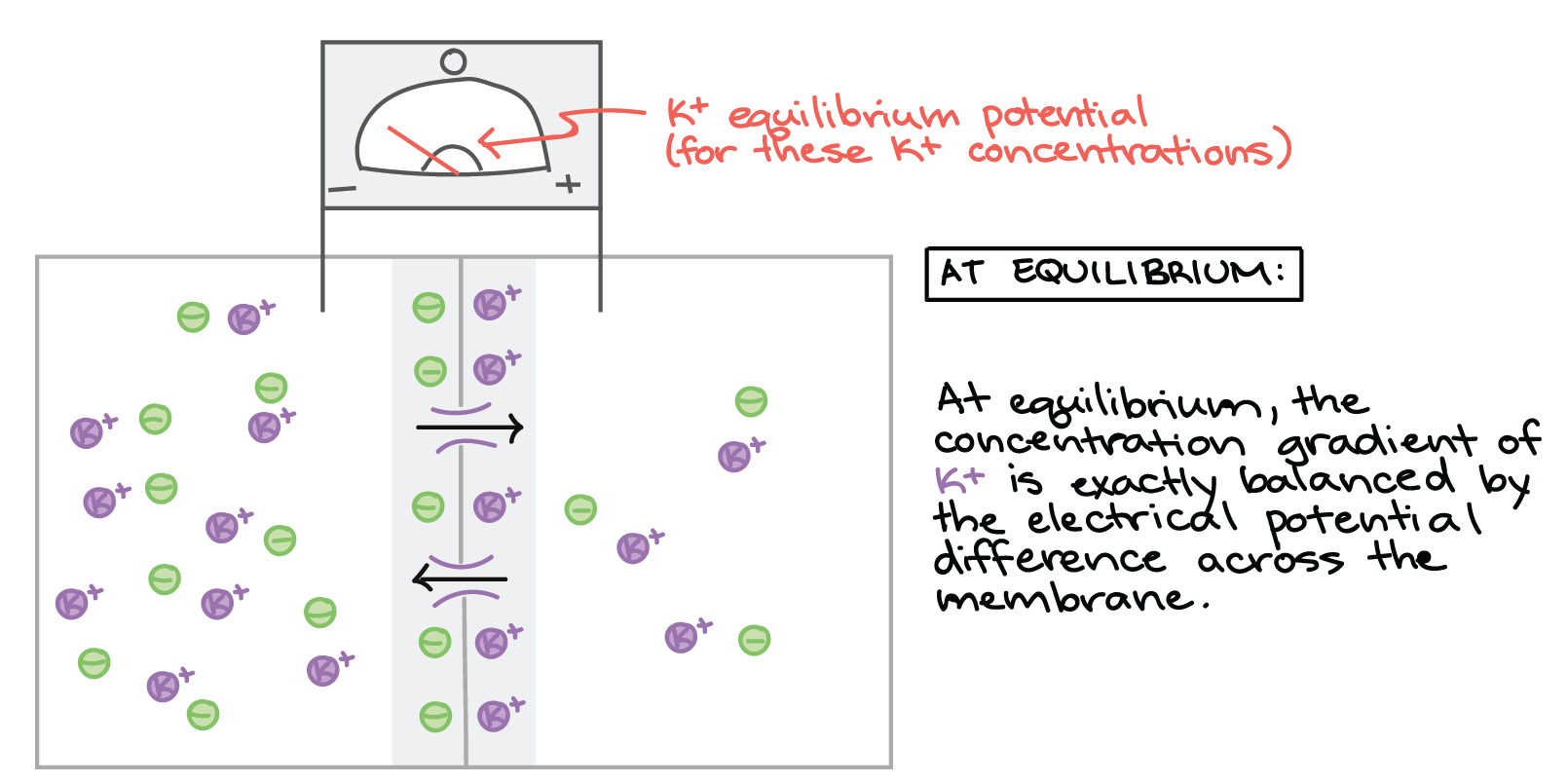
No estado de equilíbrio:
No equilíbrio, o gradiente de concentração de K+ é exatamente equilibrado pela diferença de potencial elétrico na membrana. Embora os íons K+ ainda atravessem a membrana por canais, não existe movimento de K+ de um lado para outro. O voltímetro registra um potencial de membrana negativo que é igual ao equilíbrio potencial de K+ (para concentrações de K+ presentes na célula e no fluido ao redor).
O potencial de equilíbrio
A diferença do potencial elétrico na membrana celular que equilibra exatamente o gradiente de concentração de um íon é conhecido como potencial de equilíbrio. Devido ao sistema estar em equilíbrio, o potencial da membrana tenderá a ficar em potencial de equilíbrio. Para uma célula em que existe apenas uma espécie iônica permeante (apenas um tipo de íon que consegue atravessar a membrana), o potencial de repouso da membrana será igual ao potencial de equilíbrio para esse íon.
Quanto mais acentuado é o gradiente de concentração, maior o potencial elétrico que o equilibra deve ser. Você pode obter ter uma ideia disso ao imaginar as concentrações iônicas em ambos os lados da membrana como colinas de tamanhos diferentes e pensar no potencial de equilíbrio como a força que você precisaria exercer para impedir uma pedra de rolar para baixo nas encostas.
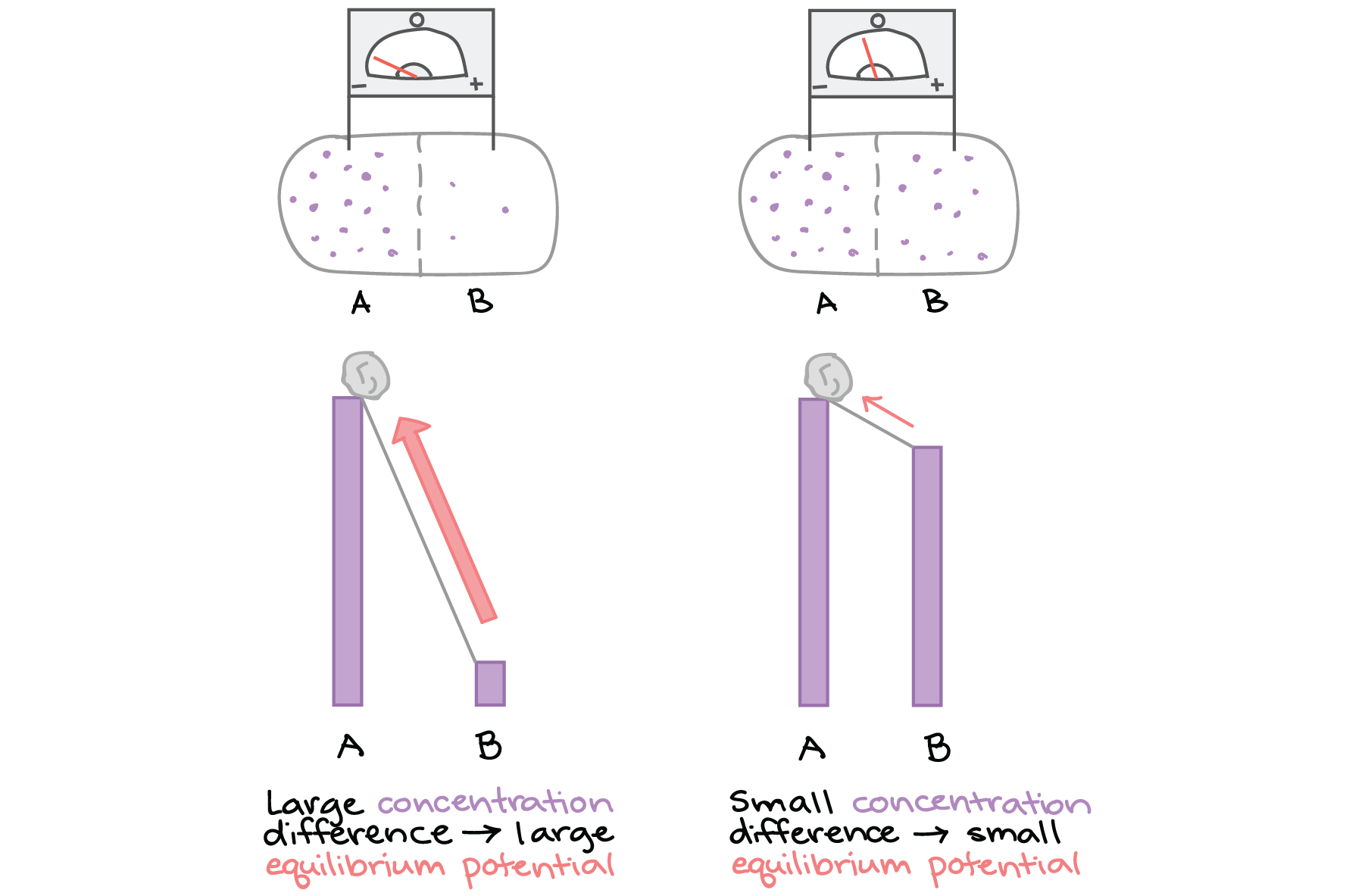
Painel esquerdo: dois compartimentos separados por uma membrana semipermeável, rotulados de A e B. Há um voltímetro entre A e B. O íon de interesse é muito mais concentrado em A do que em B, e o voltímetro com eletrodos em A e B registra uma grande tensão negativa. A tensão é análoga à força que temos que exercer para evitar que uma pedra role de um lugar muito alto de uma colina.
Painel direito: A mesma configuração, mas com o A e B apresentando uma diferença muito mais sutil na concentração do íon de interesse (B um pouco menos concentrado que A). Nesse caso, a tensão é apenas ligeiramente negativa. Isso é análogo ao caso onde temos um lugar alto e um lugar ligeiramente mais baixo e estamos exercendo uma força para impedir que uma pedra role nessa colina não tão íngreme.
Se você conhece a concentração de \text{K}^+K+K, start superscript, plus, end superscript em ambos os lados da membrana celular, então você pode prever o tamanho do potencial de equilíbrio do potássio.
O potencial de membrana é equivalente ao potencial de equilíbrio de \text K^+K+K, start superscript, plus, end superscript?
Nas células gliais, que fornecem suporte às células do sistema nervoso, o potencial de repouso da membrana é igual ao potencial de equilíbrio do \text K^+K+K, start superscript, plus, end superscript.
Nos neurônios, porém, o potencial de repouso da membrana é próximo, mas não idêntico ao potencial de equilíbrio do \text K^+K+K, start superscript, plus, end superscript. Em vez disso, em condições fisiológicas (condições como aquelas do corpo), os potenciais de repouso da membrana dos neurônios são ligeiramente menos negativos que o potencial de equilíbrio do \text K^+K+K, start superscript, plus, end superscript.
O que isso significa? Em um neurônio, outros tipos de íons além do \text K^+K+K, start superscript, plus, end superscript devem contribuir significativamente para o potencial de repouso da membrana.
[Que experiência você faria para descobrir isso?]
\text K^+K, start superscript, plus, end superscript\text {K}^+K, start superscript, plus, end superscript
\text K^+K, start superscript, plus, end superscript\text K^+K, start superscript, plus, end superscript\text K^+K, start superscript, plus, end superscript
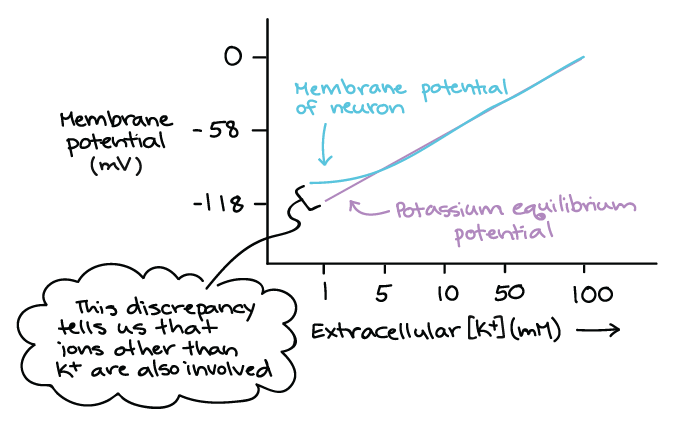
Gráfico mostrando o potencial da membrana (mV) no eixo Y e extracelular [K+] (mM) no eixo X.
O potencial de equilíbrio do potássio forma uma linha reta diagonal com a inclinação positiva deste gráfico.
O potencial de membrana real de um neurônio segue o potencial de equilíbrio do potássio na maior parte do gráfico. No entanto, ele se desvia em baixas concentrações de K + em que é mais elevado (menos negativo) do que o potencial de equilíbrio do potássio.
Esta discrepância nos diz que outros íons diferentes de K+ também estão envolvidos.
Tanto o \text K^+K+K, start superscript, plus, end superscript quanto o \text {Na}^+Na+N, a, start superscript, plus, end superscript contribuem com o potencial de repouso dos neurônios
Ao que parece, a maioria dos neurônios em repouso são permeáveis ao \text {Na}^+Na+N, a, start superscript, plus, end superscript e ao \text{Cl}^-Cl−C, l, start superscript, minus, end superscript assim como ao \text K^+K+K, start superscript, plus, end superscript. A permeabilidade ao \text{Na}^+Na+N, a, start superscript, plus, end superscript, em particular, é a principal razão pela qual o potencial de repouso da membrana é diferente do potencial de equilíbrio do potássio.
Voltemos ao nosso modelo de uma célula permeável a apenas um tipo de íon, e vamos imaginar que o \text{Na}^+Na+N, a, start superscript, plus, end superscript (em vez de \text K^+K+K, start superscript, plus, end superscript) é o único íon que pode atravessar a membrana. O \text{Na}^+Na+N, a, start superscript, plus, end superscript geralmente está presente em uma concentração muito maior fora de uma célula do que dentro, assim ele se moverá de acordo com seu gradiente de concentração para dentro da célula, fazendo com que o interior da célula fique positivo em relação ao exterior.
Devido a isso, o potencial de equilíbrio do sódio — a diferença de potencial elétrica através da membrana celular que equilibra exatamente o gradiente de concentração de \text{Na}^+Na+N, a, start superscript, plus, end superscript — será positivo. Então, em um sistema onde o \text{Na}^+Na+N, a, start superscript, plus, end superscript é o único íon permeante, o potencial da membrana será positivo.
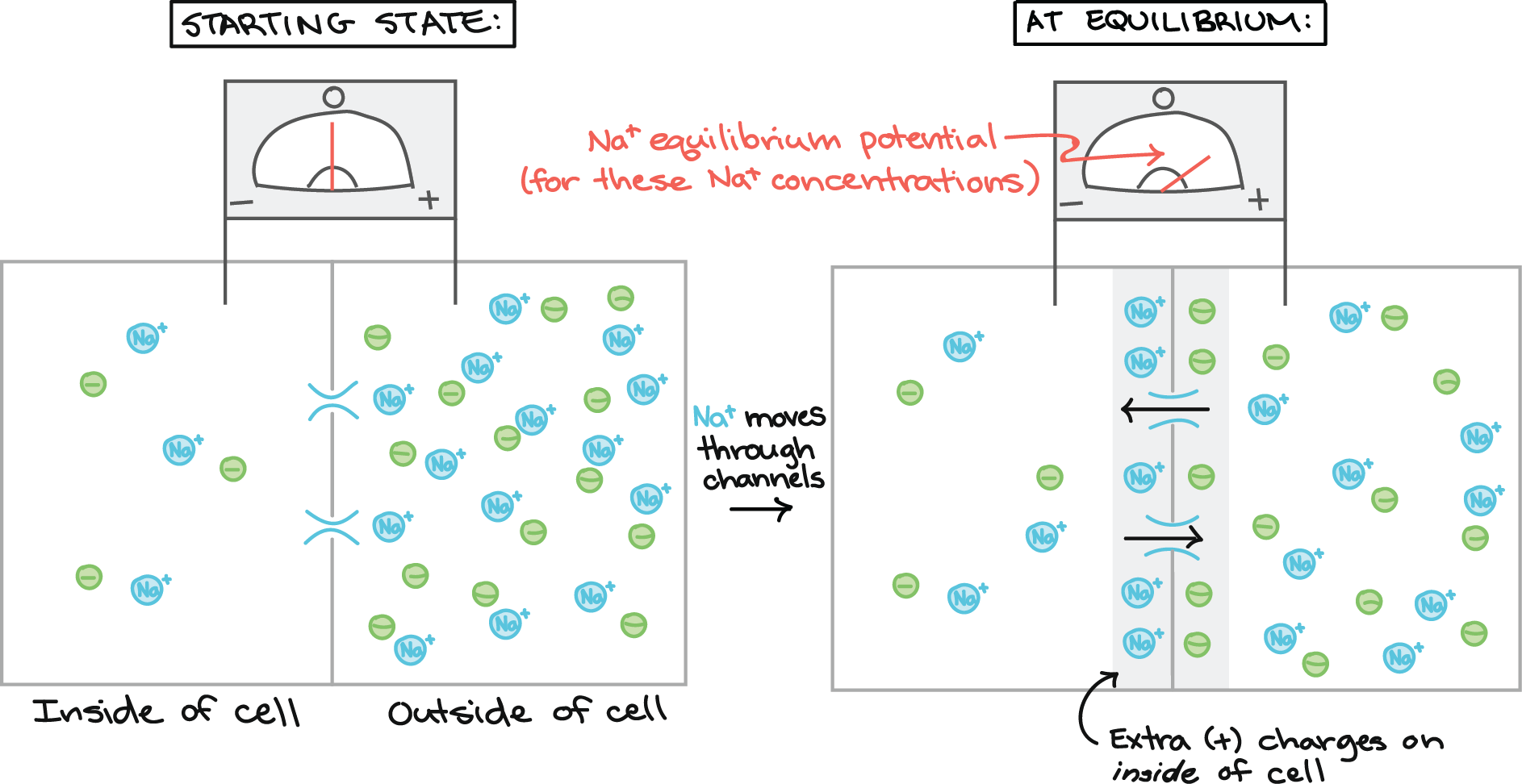
Estado inicial:
Tensão zero através da membrana, medida por um voltímetro com um eletrodo dentro e um eletrodo fora da célula. O interior da célula tem uma baixa concentração de íons de sódio, e o exterior da célula tem uma maior concentração de íons de sódio. Cada íon de sódio é contrabalançado por um ânion que se encontra do mesmo lado da membrana que o íon de sódio. Existem canais de sódio na membrana, mas eles inicialmente estão fechados.
Os canais abertos e o Na + pode se mover através deles.
No estado de equilíbrio:
O voltímetro registra agora uma tensão positiva igual ao potencial de equilíbrio do sódio para esse par de concentrações de sódio em particular. Os íons de Na+ diminuem seu gradiente de concentração até que seus movimentos se oponham por uma diferença de potencial elétrico compensatória através da membrana. Há cargas positivas extras no interior da célula na forma de íons de Na+, e esses íons de Na+ alinham-se ao longo da membrana. No lado oposto da membrana, existem ânions extras (os antigos parceiros dos íons de Na+, que são incapazes de atravessar), que também se alinham na membrana.
Em um neurônio em repouso, ambos \text{Na}^+Na+N, a, start superscript, plus, end superscript e \text{K}^+K+K, start superscript, plus, end superscript são permeantes, ou capazes de atravessar a membrana.
-
\text{Na}^+Na+N, a, start superscript, plus, end superscript vai tentar alterar o potencial da membrana em direção ao seu potencial de equilíbrio (positivo).
-
\text{K}^+K+K, start superscript, plus, end superscript vai tentar alterar o potencial da membrana em direção ao seu potencial de equilíbrio (negativo).
Você pode pensar nisso como sendo um cabo de guerra. O potencial da membrana real estará entre o potencial de equilíbrio do \text{Na}^+Na+N, a, start superscript, plus, end superscript e o potencial de equilíbrio do \text{K}^+K+K, start superscript, plus, end superscript. No entanto, o potencial da membrana real será próximo ao potencial de equilíbrio do íon com maior permeabilidade (aquele que pode atravessar a membrana mais facilmente).
[Eu gostaria de uma explicação mais técnica sobre esse conceito]
\text{Na}^+N, a, start superscript, plus, end superscript\text{K}^+K, start superscript, plus, end superscript\text {Cl} ^-C, l, start superscript, minus, end superscript
Abertura e fechamento dos canais de íons altera o potencial de membrana
Em um neurônio, o potencial de repouso da membrana está mais próximo do potencial de equilíbrio do potássio do que do potencial de equilíbrio do sódio. Isso é porque a membrana em repouso é muito mais permeável ao \text K^+K+K, start superscript, plus, end superscript do que ao \text {Na}^+Na+N, a, start superscript, plus, end superscript.
-
Se mais canais de potássio se abrirem — tornando-se ainda mais fácil para o \text{K}^+K+K, start superscript, plus, end superscript atravessar a membrana celular — a membrana iria se hiperpolarizar, ficando ainda mais perto do potencial de equilíbrio do potássio.
-
Se, por outro lado, os canais de sódio adicionais se abrirem — tornando mais fácil para o \text{Na}^+Na+N, a, start superscript, plus, end superscript atravessar a membrana — a membrana celular seria despolarizada em direção ao potencial de equilíbrio do sódio.
A alteração do número de canais iônicos abertos fornece uma maneira para controlar o potencial da membrana da célula e uma ótima maneira de produzir sinais elétricos. (Vamos ver a abertura e fechamento de canais novamente quando discutirmos sobre potenciais de ação.)
A bomba de \text{Na}^+Na+N, a, start superscript, plus, end superscript-\text K^+K+K, start superscript, plus, end superscriptmantém os gradientes para \text{Na}^+Na+N, a, start superscript, plus, end superscript e \text{K}^+K+K, start superscript, plus, end superscript
Os gradientes de concentração de \text{Na}^+Na+N, a, start superscript, plus, end superscript e \text{K}^+K+K, start superscript, plus, end superscript através da membrana da célula (e, portanto, o potencial de repouso da membrana) são mantidos pela atividade de uma proteína chamada \text{Na}^+Na+N, a, start superscript, plus, end superscript-\text K^+K+K, start superscript, plus, end superscript ATPase, muitas vezes referida como bomba de sódio-potássio. Se a bomba de \text{Na}^+Na+N, a, start superscript, plus, end superscript-\text K^+K+K, start superscript, plus, end superscriptfor desligada, os gradientes de concentração \text{Na}^+Na+N, a, start superscript, plus, end superscript e \text K^+K+K, start superscript, plus, end superscript se dissiparão e o potencial da membrana também.
[Por que é necessário uma bomba para manter os gradientes de concentração?]
\text {Na} ^+N, a, start superscript, plus, end superscript\text K^+K, start superscript, plus, end superscript\text K^+K, start superscript, plus, end superscript\text {Na}^+N, a, start superscript, plus, end superscript\text K^+K, start superscript, plus, end superscript\text {Na}^+N, a, start superscript, plus, end superscript
\text{Na}^+N, a, start superscript, plus, end superscript\text K^+K, start superscript, plus, end superscript
Assim como os canais de íons que permitem que \text{Na}^+Na+N, a, start superscript, plus, end superscript e \text K^+K+K, start superscript, plus, end superscript atravessem a membrana celular, a bomba \text{Na}^+Na+N, a, start superscript, plus, end superscript-\text K^+K+K, start superscript, plus, end superscript é uma proteína que abrange a membrana. Ao contrário dos canais de potássio e de sódio, no entanto, a bomba de \text{Na}^+Na+N, a, start superscript, plus, end superscript-\text K^+K+K, start superscript, plus, end superscript não fornece apenas uma direção para que \text {Na}^+Na+N, a, start superscript, plus, end superscript e \text K^+K+K, start superscript, plus, end superscript movimentem seus gradientes eletroquímicos. Em vez disso, ela transporta ativamente \text{Na}^+Na+N, a, start superscript, plus, end superscript e \text{K}^+K+K, start superscript, plus, end superscript contra seus gradientes eletroquímicos.
A energia para esse movimento "para cima" vem da hidrólise do ATP (a divisão do ATP em ADP e fosfato inorgânico). Para cada molécula de ATP que é quebrada, 333 íons de \text{Na}^+Na+N, a, start superscript, plus, end superscript são movidos do interior para o exterior da célula, e 222 íons de \text K^+K+K, start superscript, plus, end superscript são movidos do exterior para o interior.
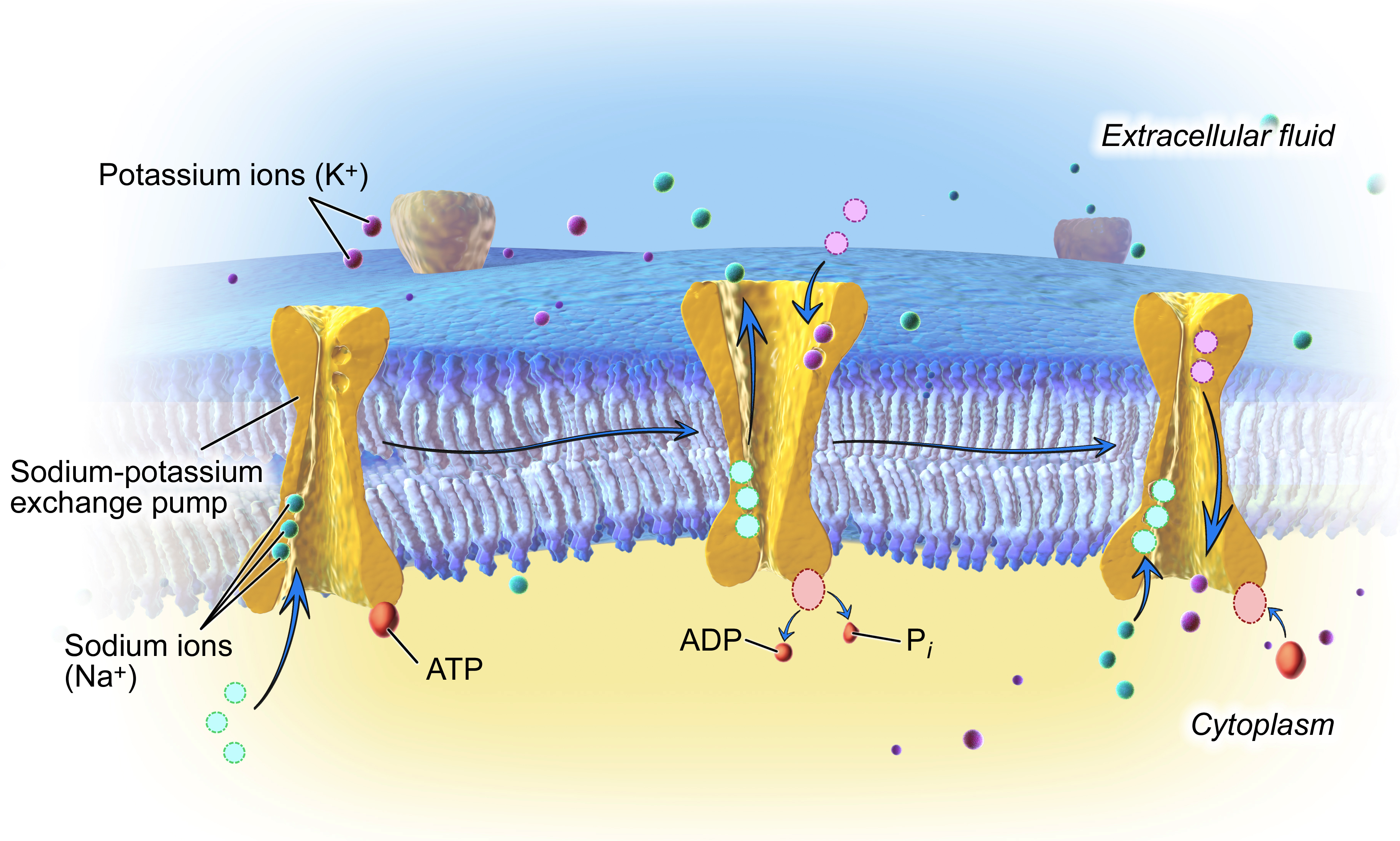
-
Três íons de sódio ligam-se a bomba de potássio de sódio –, que está aberta para o interior da célula.
-
A bomba hidrolisa ATP, fosforilando-se (anexando um grupo fosfato a si) e liberando ATP. Este evento de fosforilação provoca uma mudança na forma na bomba, na qual ela se fecha do lado voltado para o interior da célula e abre-se para o exterior da célula. Os três íons de sódio são liberados e dois íons de potássio ligam-se no interior da bomba.
-
A ligação dos íons potássio aciona outra mudança de forma na bomba, que perde seu grupo fosfato e retorna à sua forma voltada para o interior. Os íons de potássio são liberados para o interior da célula, e o ciclo da bomba pode recomeçar.
_Imagem modificada de"The sodium-potassium exchange pump," pelo staff de Blausen (CC BY 3,0)._
[Veja detalhes de como a bomba transporta íons]
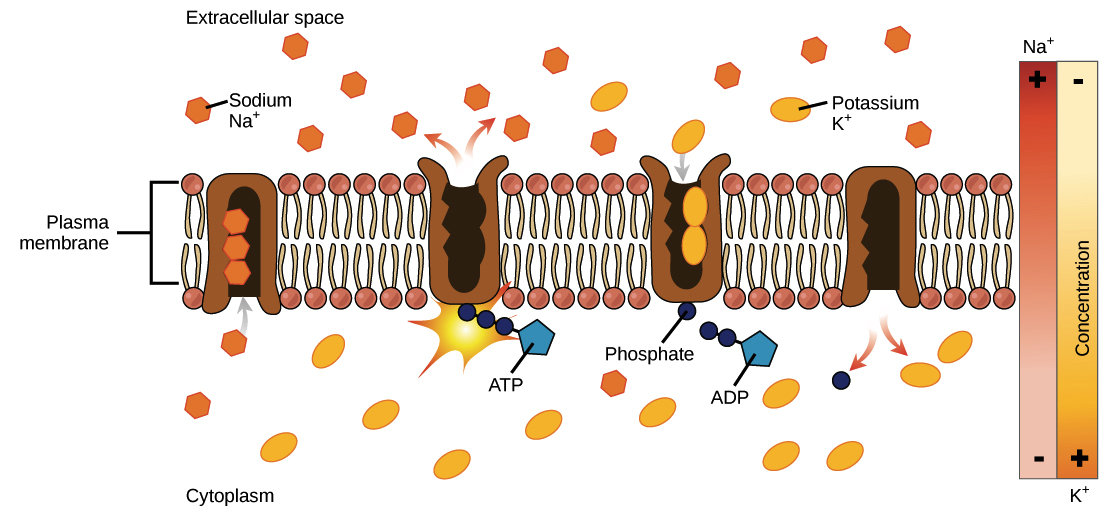
-
No início, a bomba está aberta para o interior da célula. Nessa forma, a bomba liga-se prontamente aos íons de \text {Na}^+Na+N, a, start superscript, plus, end superscript (tem uma grande afinidade com eles) e vai usar três deles.
-
Quando os íons de \text {Na}^+Na+N, a, start superscript, plus, end superscript se ligam, disparam a bomba para a hidrólise (quebra) do ATP. Um grupo fosfato do ATP é anexado à bomba, que é então chamada de fosforilada. O ADP é liberado como um subproduto.
-
A fosforilação faz a bomba mudar de forma, reorientando-se para abrir na direção do espaço extracelular. Nessa conformação, a bomba deixa de se ligar aos íons de \text {Na}^+Na+N, a, start superscript, plus, end superscript (fica com baixa afinidade com eles), e então três íons de \text {Na}^+Na+N, a, start superscript, plus, end superscript são liberados para o exterior da célula.
-
Em sua forma voltada para fora, a bomba muda de constância e agora se liga prontamente (tem alta afinidade) aos íons de \text K^+K+K, start superscript, plus, end superscript. Ela vai se ligar a dois deles, e isto aciona a remoção do grupo fosfato ligado à bomba na etapa 2.
-
Com a saída do grupo fosfato, a bomba vai mudar de volta para sua forma original, abrindo-se em direção ao interior da célula.
-
Em sua forma voltada para dentro, a bomba não mais prontamente se vincula aos íons de \text {K}^+K+K, start superscript, plus, end superscript, então os dois íons de \text {K}^+K+K, start superscript, plus, end superscript serão liberados para o citoplasma. A bomba, agora, volta para onde estava na etapa 1, e o ciclo pode começar outra vez.
-
\text {Na}^+N, a, start superscript, plus, end superscript
-
\text {Na}^+N, a, start superscript, plus, end superscript
-
\text {Na}^+N, a, start superscript, plus, end superscript\text {Na}^+N, a, start superscript, plus, end superscript
-
\text K^+K, start superscript, plus, end superscript
-
\text {K}^+K, start superscript, plus, end superscript\text {K}^+K, start superscript, plus, end superscript
Como os 333 \text{Na}^+Na+N, a, start superscript, plus, end superscript são exportados para cada 222 \text K^+K+K, start superscript, plus, end superscript trazidos para o interior da célula, a bomba faz uma pequena contribuição direta com o potencial de repouso da membrana (tornando-a um pouco mais negativa do que seria). A grande contribuição da bomba para o potencial da membrana, no entanto, é indireta: ela mantém os gradientes de \text {Na}^+Na+N, a, start superscript, plus, end superscript e \text K^+K+K, start superscript, plus, end superscript estáveis, os quais dão origem ao potencial da membrana à medida que \text {Na}^+Na+N, a, start superscript, plus, end superscript e \text K^+K+K, start superscript, plus, end superscript diminuem seus respectivos gradientes de concentração através de canais de vazamento.
Andre Smaira
O potencial de ação é caracterizado por uma forte e repentina alteração do potencial de membrana. Durante o potencial de repouso a maioria dos canais iônicos estão fechados, não permitindo a passagem de íons entre o meio intra e extracelular. Quando, por exemplo, por um input sináptico, há uma depolarização local da membrana, os íons positivos passam através de canais iônicos para o interior da célula. Com a entrada dos íons positivos o interior se torna menos negativo. Quando esse fluxo de íons é suficiente, ele causa a abertura de canais dependentes dessa diferença de potencial, levando a uma forte depolarização e fazendo com que o interior da célula se torne positivo em relação ao exterior da célula, causando um potencial de ação. A depolarização da membrana que leva a um potencial de ação é chamado de estímulo.
Em especial, o potencial de ação, também chamado de potencial de equilíbrio, é o movimento de íons dentro e fora da célula. Ele envolve especificamente íons sódio e potássio. Eles são movidos para dentro e para fora da célula através de canais de sódio e potássio e bomba de sódio-potássio.
Existem cinco fases de potencial de ação cardíaco, numeradas de 0 a 4.
- A fase 0 é a despolarização da membrana e a abertura de canais de sódio "rápidos" (ou seja, de alto fluxo). O fluxo de potássio também diminui. A despolarização acontece em razão do significativo aumento na permeabilidade aos íons sódio na membrana celular, que, consequentemente, acarreta em um grande fluxo de íons sódio para dentro da célula através do processo de difusão simples;
- A fase 1 é a repolarização parcial da membrana, graças a uma rápida diminuição na passagem de íons de sódio à medida que os canais de sódio se fecham rapidamente;
- A fase 2 é a fase de platô, na qual o movimento de íons de cálcio para fora da célula mantém a despolarização. Recebe este nome porque a carga elétrica através da membrana muda muito pouco nesta fase;
- A fase 3 é a repolarização, quando os canais de sódio e cálcio se fecham e o potencial de membrana retorna ao seu nível basal;
- A fase 4 vê a membrana em seu chamado potencial de repouso de -90 milivolts (mV) como resultado do trabalho da bomba de íon \(\require{text}\text{Na}^+\)/\(\require{text}\text{K}^{+}\)valor é negativo porque o potencial dentro da célula é negativo comparado ao potencial externo, e o último é tratado como o quadro de referência zero.
Mais especificamente, o potencial de repouso acontece devido a diferença de concentração entre os íons de sódio \((\require{text}\text{Na}^{+})\)e potássio \((\require{text}\text{K}^{+})\)no interior e no exterior da célula, que são mantidos pelo bombeamento ativo de tais íons pelas membranas celulares, que fazem com que o sódio saia e o potássio entre na célula.

Faça como milhares de estudantes: teste grátis o Passei Direto

Esse e outros conteúdos desbloqueados

16 milhões de materiais de várias disciplinas

Impressão de materiais
Agora você pode testar o
Passei Direto grátis
✏️ Responder
Para escrever sua resposta aqui, entre ou crie uma conta
Outros materiais
Perguntas relacionadas
qual a diferença de intracelular e extracelular e onde são encontrados corpo?
Fisiologia Humana I
•ESTÁCIO EAD
Ana Lúcia Sousa
Qual a diferença entre um líquido intracelular e um líquido extracelular?
Morfologia e Fisiologia Humana
•UNISC
MARCELO AZEVEDO


{kind=link}
Compartilhar